![]()
Research Article
Neuroscience
Development of auditory and spontaneous movement responses to music over the first postnatal year
Trinh Nguyen
![]()
Félix Bigand
Susanne Reisner
Atesh Koul
Roberta Bianco
Gabriela Markova
Stefanie Hoehl
Giacomo Novembre
![]()
Neuroscience of Perception and Action Lab, Italian Institute of Technology, Italy;
Department of Developmental and Educational Psychology, University of Vienna, Austria;
Department of Developmental and Biological Psychology, Heidelberg University, Germany;
Doctoral School Cognition, Behavior and Neuroscience, University of Vienna, Austria;
Department of Translational Research and New Technologies in Medicine and Surgery, University of Pisa, Italy;
Institute for Early Life Care, Paracelsus Medical University, Austria;
https://doi.org/10.7554/eLife.107088.4
Open access
Copyright information
Download
Cite
Share
Comment Open annotations (there are currently 0 annotations on this page).
Comment Open annotations (there are currently 0 annotations on this page).
1,033 views
73 downloads
2 citations
Altmetric provides a collated score for online attention across various platforms and media.
See more details
Bluesky Streamline Icon: https://streamlinehq.com
Bluesky
[Bluesky Streamline Icon: https://streamlinehq.com
Bluesky](https://bsky.app/intent/compose?text=Development%20of%20auditory%20and%20spontaneous%20movement%20responses%20to%20music%20over%20the%20first%20postnatal%20year%20https%3A%2F%2Fdoi.org%2F10.7554%2FeLife.107088.4)
Threads Fill Streamline Icon: https://streamlinehq.com
Threads Fill Streamline Icon: https://streamlinehq.com
Trinh Nguyen
Félix Bigand
Susanne Reisner
Atesh Koul
Roberta Bianco
Gabriela Markova
Stefanie Hoehl
Giacomo Novembre
Copy to clipboard
Download BibTeX
Download .RIS
eLife Assessment
This well-designed study offers important insights into the development of infants' responses to music based on the exploration of EEG neural auditory responses and video-based movement analysis. The compelling results revealed that evoked responses emerge between 3 and 12 months of age, but no age group demonstrated evidence of coordinated movements to music. This study will be of significant interest to developmental psychologists and neuroscientists, as well as researchers interested in music processing and in the translation of perception into action.
https://doi.org/10.7554/eLife.107088.4.sa0
Important: Findings that have theoretical or practical implications beyond a single subfield
Landmark
Fundamental
Important
Valuable
Useful
Compelling: Evidence that features methods, data and analyses more rigorous than the current state-of-the-art
Exceptional
Compelling
Convincing
Solid
Incomplete
Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Learn more about eLife Assessments
Abstract
eLife digest
Introduction
Results
Discussion
Materials and methods
Appendix 1
Appendix 2
Data availability
References
Article and author information
Article and author information
Metrics
Abstract
Humans across cultures not only share the ability to recognise music but also respond to it through movement. While the sensory encoding of music is well-studied, when and how infants naturally start moving to music is largely unexplored. This study simultaneously investigates infants’ neural (auditory) responses and spontaneous movements to music during the first postnatal year. Neural activity (EEG) and body kinematics (markerless pose estimation) were recorded from 79 infants (aged 3, 6, and 12 months) listening to refrains of children’s music, along with shuffled, high-pitched, and low-pitched versions of the same songs. Neural data revealed that, across all ages, infants exhibit enhanced auditory responses to music compared to shuffled music, indicating that auditory encoding of music emerges early in development. Movement data revealed a different outcome. While coarse auditory-motor coupling is present at all ages, more complex structured movement patterns emerge in response to music only by 12 months. Notably, no age group demonstrated evidence of coordinated movements to music. Additionally, enhanced auditory responses to high vs low pitch were only evident at 6 months, while infants’ movements were better predicted by high-pitched compared to low-pitched music at all ages. This study provides initial insights into how the developing brain gradually transforms music into spontaneous movements of increasing complexity.
eLife digest
Most people, no matter where they grow up, enjoy listening to music – and many instinctively move their bodies to it. This universal behavior raises a fascinating question: when does the brain first respond to music, and how does that ability develop?
Babies are born with a natural sensitivity to sound. Their brains can already detect patterns in what they hear, such as repeated rhythms and melodies. Scientists can measure this brain activity using an EEG (electroencephalography), which records electrical signals produced by the brain in response to sounds. Infants also naturally move their bodies in response to sounds around them. However, we do not fully understand when these two abilities – recognizing music and moving to it – emerge, or how they relate during the first year of life.
Nguyen et al. wanted to understand how babies' brain responses to music and their spontaneous body movements to music develop during the first year of life. The researchers also asked whether pitch – high or low music sounds – affects these two responses differently, since babies are known to be drawn to high-pitched sounds.
Nguyen et al. tested 79 infants aged 3, 6, and 12 months by playing children's songs and scrambled versions of the same songs. They measured brain activity using electroencephalography (EEG) while also tracking and reconstructing full-body movements from video recordings.
The results revealed that all age groups – even 3-month-olds – showed stronger brain responses to real music than to scrambled music, indicating that the brain encodes musical structure very early in life. However, only 12-month-olds spontaneously moved more to music than to scrambled music, specifically exhibiting rocking, swaying, and clapping-like movements. Importantly, no age group showed movements that were coordinated in time with the musical beat. Additionally, only 6-month-olds showed stronger brain responses to high-pitched compared to low-pitched music, while high-pitched music predicted movements at all ages.
Nguyen et al. are the first to measure both brain activity and body movement simultaneously in infants this young. Their findings will be relevant to researchers studying how children develop musical and movement skills, and how early rhythmic responses eventually give rise to dancing. They also provide valuable insights for caregivers and early childhood educators who use music to engage and support infants. Before any practical applications can be developed, future studies should examine how music-driven movement coordination continues to develop beyond 12 months and investigate the brain pathways that link hearing music to moving – and eventually dancing – to it.
Introduction
Musicality – the biological predisposition to perceive, appreciate, and produce music (Honing, 2018; Trehub, 2003) – is increasingly recognised as a fundamental aspect of human nature. Numerous accounts suggest that engaging with music through movement is at the core of musicality (Honing et al., 2015; Schachner et al., 2009; Trehub et al., 2015). Functionally, such engagement can be broken down into two fundamental components of neurocognitive development: the ability to perceive and recognise music (sensory component), and the ability to produce movement responses that are temporally aligned with the musical structure, from coordinated vocalizations and percussive actions up to complex dance moves (motor component; Brown, 2022; Trehub, 2003; Trevarthen, 1999). Despite this inherent predisposition toward music, the developmental trajectory of infants’ musicality remains largely unknown (see Nguyen et al., 2023a, for a review). While there is increasing research on infant music perception, including controlled manipulations of select musical features, we know less about the translation of perception into action, namely the ontogenesis of infants’ spontaneous movements to music (see Fujii et al., 2014; Nguyen et al., 2023b; Zentner and Eerola, 2010). Furthermore, making our understanding of music-driven motor engagement even more incomplete, no studies to date have looked at both brain activity and spontaneous body movements simultaneously, especially during the first year of life. Accordingly, how the processing of music and its features is transformed into organised motor responses remains underexplored.
The sensory component of musicality, namely music perception, can be measured using electroencephalography (EEG), specifically by recording cortical auditory evoked potentials (event-related potentials [ERP]). One of these responses is the infantile P1, a phase-locked EEG positivity peaking around 200-300 ms after an auditory stimulus (Chen et al., 2016; Kushnerenko et al., 2002; Wunderlich et al., 2006). The infantile P1 has been observed in response to both musical notes and speech segments. Auditory evoked potentials, when elicited isochronously, can also be captured using frequency domain analyses (Damsma et al., 2024; Novembre and Iannetti, 2018), such as auditory steady-state responses (ASSR), which are also called steady-state evoked potentials (SSEP, e.g. Cirelli et al., 2016; Nave et al., 2022). These neural responses can provide insight into the developing auditory system and its ability to encode musical structure. Using these neurophysiological measures, prior research has shown that newborns and infants are sensitive to beat structure, pitch deviants, and tone interval regularities (Bianco et al., 2025; Edalati et al., 2023; Háden et al., 2022; Háden et al., 2015; Háden et al., 2009; Stefanics et al., 2009; Winkler et al., 2009). Despite these promising results, the neurophysiology of early music processing – particularly its developmental trajectory – remains not fully understood. Here, our primary goal is to investigate infants’ neural encoding of music utilizing both ERP and ASSR approaches to characterize how such neural responses change across the first year of life.
Another component of musicality is the capacity to move to music (motor component; Brown, 2022; Fitch, 2015; Honing et al., 2015; Trehub et al., 2015). This capacity is linked to infants not only recognising musical structure but also moving their bodies in response to it. Even though this capacity appears to develop precociously, as evidenced by the fact that even 28-35 week-old foetuses move to music (Kisilevsky et al., 2004), very few studies have systematically examined music-driven spontaneous body movements in infants. An influential paper by Zentner and Eerola, 2010 reported that infants across a large age range (from 5 to 24 months) showed more spontaneous rhythmic movements in response to classical music and children’s music compared to infant-directed speech. Importantly, their movements were not synchronized with the musical input, even though a small degree of tempo flexibility was observed (i.e. faster musical tempi evoked relatively faster movement periodicities). The lack of synchrony between music and body movements has also been reported in younger (i.e. 3-4 months old) infants listening to popular music (Fujii et al., 2014). Furthermore, another study testing 7-month-old infants reported more movement in response to (sung) playsongs compared to lullabies but did not assess movement synchrony (Nguyen et al., 2023b). Despite these initial investigations, it remains unclear when infants begin to move in response to music, which specific movements are evoked, and when these movements become coordinated with the music. Moreover, a critical limitation in existing research is the lack of a control condition to determine whether these movements are driven specifically by musical structure or reflect general motor activity in response to auditory input. As a second goal, this study is the first to systematically test the gradual development of music-induced movements in different age groups across the first year of life.
Music engages both sensory and motor systems, yet different musical features may differentially shape infants’ engagement with music. While rhythm has been widely studied in early music cognition, pitch is another salient acoustic cue that could play a role in auditory-motor engagement, particularly in infancy. High pitch is a defining feature of infant-directed speech (Fernald and Simon, 1984), among other features, such as exaggerated intonation, slower tempo, and simplified vocabulary (Fernald and Kuhl, 1987; Kuhl and Meltzoff, 1982). Similarly, infants most frequently listen to music characterized by high pitch (Costa-Giomi and Sun, 2016; Nakata and Trehub, 2011). Reflecting its prominence, high pitch is found to be one of the most prominent features thought to effectively capture (Conrad et al., 2011; Eckerdal and Merker, 2009; Trainor, 1996; Trainor and Zacharias, 1998) and guide infants' attention (Lense et al., 2022; Trainor and Desjardins, 2002). On the neural level, infants are also better at encoding pitch deviances in the high voice of polyphonic music, thus showing high voice superiority from 3 months of age (Marie and Trainor, 2013; Marie and Trainor, 2014). Taken together, these findings indicate that higher-pitch music would amplify infants’ neural responses (i.e. sensory component) in comparison to lower-pitch music. On the other hand, we know that adults move more to music with greater energy in lower frequencies (Cameron et al., 2022; Stupacher et al., 2013; Stupacher et al., 2016; Van Dyck et al., 2013). Yet, it remains unknown whether low-pitch music elicits increased movement in infants, as it does in adults, or whether infants’ attraction to high pitch also extends to enhance their motor responses. As a third goal, we thus investigate how musical pitch affects infants’ sensory and motor components.
We presented infants, aged 3, 6, and 12 months, with instrumental refrains of children’s songs (music), shuffled versions of the same songs (shuffled music), and transpositions of the songs that would either emphasize the melody (high pitch) or the bassline (low pitch). We recorded infants’ neural activity using EEG and specifically extracted ERPs and ASSR as indices of infants’ neural response to the various auditory stimuli. We also analysed spontaneous (full-body) movement kinematics using automated video-based motion tracking (DeepLabCut) and extracted principal movements using principal component analysis (see Figure 1, c.f. Bigand et al., 2024a; Toiviainen et al., 2010). By adopting a cross-sectional design, we aimed to characterize the maturation of both auditory and movement responses across infancy. We hypothesized that auditory responses would be enhanced when triggered by music compared to shuffled music. This hypothesis was based on the notion that musical structure, notably eroded in the shuffled musical stimuli, is essential to attract infants’ attention towards predictable events (Kouider et al., 2015; Lense et al., 2022). Similarly, based on previous evidence comparing movement responses to music vs speech and silence (Fujii et al., 2014; Zentner and Eerola, 2010), we expected the presence of musical structure to increase the likelihood of spontaneous movements in response to music compared to shuffled music, but we did not have a specific hypothesis about which particular movements would be produced. We further hypothesized that infants would show enhanced neural responses to high- compared to low-pitch music and explored co-occurring differences in spontaneous movements. Generally, we aimed to characterize the maturation of both auditory and motor responses as infants get older. By studying both sensory and motor components of musicality, we aimed to deepen our understanding of when and how infants learn to transform what they perceive into spontaneous movements, eventually leading to the emergence of synchronization to music (Brown, 2022; Fitch, 2015; Honing et al., 2015; Patel and Iversen, 2014).
{kind=link}
{kind=link}
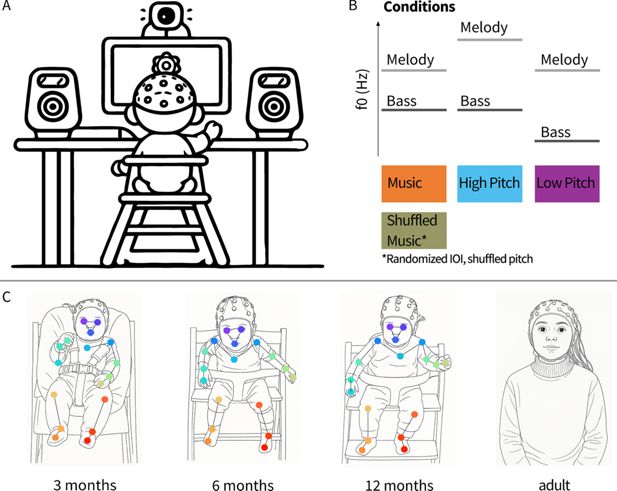
(A) Infants sat in front of a screen with speakers on each side. The screen showed slowly blossoming flowers to attract infants’ attention. Caregivers (not shown) sat behind the infants and wore noise-cancelling headphones. (B) Infants listened to polyphonic auditory stimuli consisting of a melody and a bassline in four different conditions. The music condition included two children’s songs. The shuffled music condition included versions of the songs used in the music condition that were shuffled in pitch order and randomised in inter-onset intervals (IOI). Stimuli belonging to the music and shuffled music conditions had the same set of pitches (pitch range), differing only in sequence and timing. In the high-pitch condition, the melody was shifted one octave higher than in the music condition. In the low-pitch condition, the bassline was shifted one octave lower than in the music condition. Hence, the two voices composing the high-pitch condition were one octave higher than those composing the low-pitch condition. (C) The sample included infants at 3 months (N=26), 6 months (N=26), 12 months (N=27), and an adult control sample (N=26). The dots overlaying the images represent the body parts whose movements were tracked using video-based kinematic analysis.
Results
This study included EEG and movement measurements taken from 79 infants in the first postnatal year, as well as EEG measurements taken from a control sample of 26 adults. Participants were exposed to two polyphonic children’s songs featuring a melody and a bassline, and their manipulated versions (see Table 1 and Supplementary file 1 for an acoustic characterization of the musical stimuli). We investigated neural responses and movement responses to music vs shuffled music (a control condition in which we shuffled the melody and randomized the inter-onset intervals (IOI) of the music). Additionally, we contrasted neural and movement responses to high- vs low-pitch music. Music vs shuffle conditions (manipulation of structure but not pitch height) and high pitch vs low pitch conditions (manipulation of pitch height but not structure) are contrasted separately to avoid comparing conditions differing in more than one variable. Below, we begin by reporting the neural results, followed by the movement results.
Note. M=mean, SD = standard deviation, IOI = inter-onset interval, IPI = inter peak interval, Hz = hertz (frequency). IOI and IPI were computed across melody and bass notes. Hopp refers to the Hungarian playsong (‘Hopp Juliska’), Lola refers to the Spanish playsong (‘La vaca lola’).
Note. M=mean, SD = standard deviation, IOI = inter-onset interval, IPI = inter peak interval, Hz = hertz (frequency). IOI and IPI were computed across melody and bass notes. Hopp refers to the Hungarian playsong (‘Hopp Juliska’), Lola refers to the Spanish playsong (‘La vaca lola’).
EEG: Event-related potentials (ERP)
To examine the neural responses, we analysed the average ERPs time-locked to the acoustic tone onsets of the bassline notes of the auditory stimuli (Figure 2). Adults’ ERPs, which served as a benchmark to interpret infants’ responses, included an early positivity peaking at 37 ms post-stimulus (so-called ‘P50,’ here reaching an average amplitude of 1.05 µV), followed by a later negativity peaking at 87 ms post-stimulus (so-called ‘N100,’ here reaching an amplitude of -0.43 µV) and a second positivity peaking at 158 ms post-stimulus (so-called ‘P200,’ here reaching an amplitude of 0.85 µV). This triphasic EEG pattern has been widely observed in adults in response to fast-rising auditory stimuli and across different contexts (Novembre et al., 2018; Pratt et al., 2008; Remijn et al., 2014; Somervail et al., 2021). Cluster-based permutation analyses, contrasting the ERPs elicited by music vs shuffled music, revealed that the amplitude of the P50 – hereafter referred to as P1 – was larger in response to music compared to shuffled music, particularly within the time range comprised between -17 and 58 ms post-stimulus (cluster-t=366.16, p=0.016). Similarly, the amplitude of the following P200 - hereafter referred to as P2 – was larger in response to music than to shuffled music, particularly within the time range of 114-190 ms post-stimulus (cluster-t=395.42, p=0.016). Both P1 and P2 responses were observed over frontocentral electrodes, showing a medial distribution, in line with previous literature (e.g. Lijffijt et al., 2009).
{kind=link}
{kind=link}
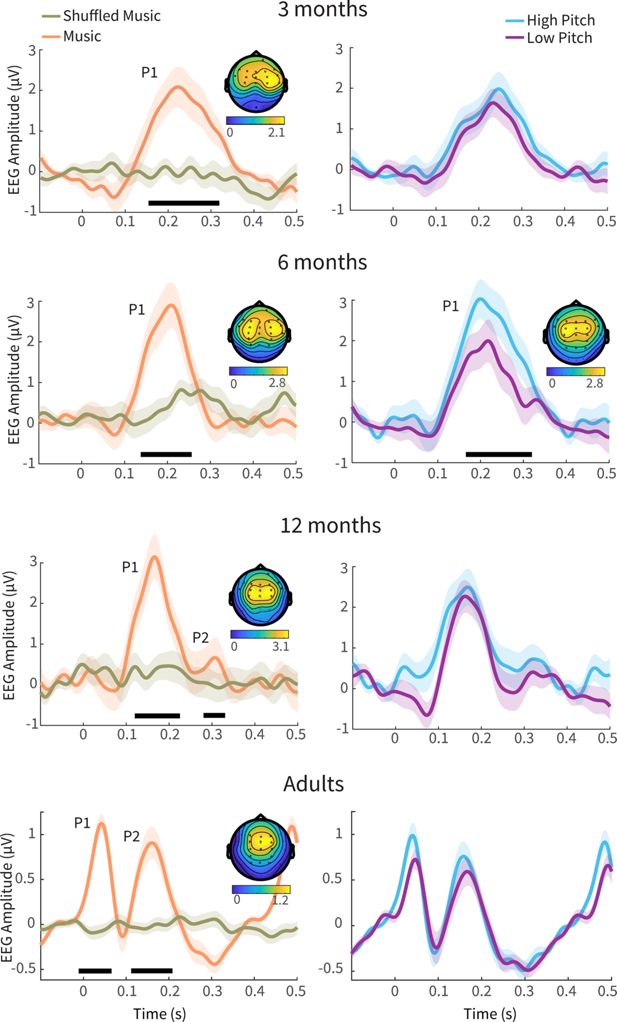
Event-related potentials (ERPs) elicited by the notes within the music (orange, left) vs shuffled music (khaki, left) as well as by the notes comprised within the high-pitch (blue, right) vs low-pitch music (purple, right), across four groups of participants (plotted in ascending order of age, from top to bottom): 3-, 6-, 12-month-old infants (N=79) and adults (N=26). Grand-average ERPs are averaged across electrodes within the significant cluster of each age group in the music condition (except for pitch condition comparison in the 6-month-olds, which used the cluster from that contrast). Shaded areas indicate the standard error. ERPs show progressively shorter latencies with increasing age. All groups exhibited a P1 response, while only older infants (12-month-olds) and adults additionally exhibited a P2. Music elicited a larger P1 (and, when present, P2) amplitude compared to shuffled music, notably across all groups (time ranges associated with a significant difference are indicated by horizontal black lines). The topography of this neural response (averaged across the time window of the P1 cluster) in the music condition appears to shift more medially with increasing age. Colorbars beneath topography plots index EEG amplitude values.
All infants’ ERPs showed a P1 response, while a P2 response was observed only in 12-month-old infants, albeit with a lower amplitude than the P1. The P1 latency decreased (χ²(2)=391.25, p<0.001), and its amplitude increased (χ²(2)=8.59, p=0.014) with age (Figure 2, left column). Importantly, and in line with the adults’ data, all infant groups exhibited enhanced P1 amplitudes in response to music compared to shuffled music. Actually, across all groups, shuffled music did not elicit as clear ERPs as the ones elicited by music. Cluster-based permutation (nPerm = 1000) testing revealed that 3-month-old infants’ P1 amplitude was enhanced between 177 and 305 ms post-stimulus (cluster-t=1111.90, p=0.002). Within this window, the P1 peaked at 212 ms and reached an average amplitude of 1.8 µV. The topography included a frontocentral cluster with a slight right lateralization. In 6-month-old infants, the amplitude of the P1 was enhanced between 116 and 284 ms post-stimulus (cluster-t=1401.60, p=0.002), peaking at 165 ms and reaching an amplitude of 2.8 µV. The topography included a few (centro-) parietal electrodes in addition to several frontocentral electrodes, with a bilateral activation. In 12-month-old infants, the amplitude of the neural response to music was enhanced in a two-peak cluster (cluster-t=1416.30, p=0.002). The first peak, an infantile P1, occurred between 104 and 227 ms, peaked at 146 ms post-stimulus, and reached an amplitude of 3.1 µV. Notably, 12-month-old infants exhibited an additional positivity, namely an infantile P2, possibly homologous to the P200 observed in adults. The P2 ranged from 307 to 325 ms post-stimulus and peaked at 316 ms, with an average amplitude of 1.0 µV. The topographies remained frontocentral but were more medial, similar to adults.
To rule out the possibility that increased neural responses in the music condition reflected differences in inter-tone temporal spacing (see IOI range in Table 1), we performed control analyses, including only epochs from the shuffled music condition for which the prior or subsequent IOI exceeded the median IOI duration. The resulting ERPs in the shuffled music condition were highly similar to those reported above (compare Figure 2 with Figure 2—figure supplement 1), and the difference between the music and shuffled music conditions remained significant across all age groups (p<0.05). These findings indicate that the observed effects are not solely attributable to variations in inter-tone spacing but also reflect sensitivity to musical structure.
Next, we examined neural responses to the notes in the high- and low-pitch conditions (Figure 2, right column). The morphology of both adults’ and infants’ ERPs was generally comparable to that elicited by the music condition. Cluster-based permutation (nPerm = 1000) testing revealed that the amplitude of the adults’ ERPs was comparable across high- and low-pitch conditions (p>0.050). This was also the case for both 3- and 12-month-old infants, but notably not for 6-month-old infants, who exhibited an enhanced P1 in response to high-pitch vs low-pitch conditions (cluster-t=763.84, p=0.002). This enhanced positivity between 178 and 332 ms peaked at 204 ms and reached an average amplitude of 2.8 µV. Similar to the neural response elicited by the music condition, the topography included few (centro-) parietal electrodes in addition to several frontocentral electrodes, with a bilateral activation.
EEG: Auditory steady state responses (ASSR)
Figure 3 shows bar plots indexing the relative power of ASSRs elicited by the auditory stimuli (power estimates were averaged across the electrodes comprised within the ERP clusters that were common to all age groups, i.e. FP2, F7, F3, Fz, F4, F8, FC7, FC3, FCz, FC4, FC8, C3, Cz, C4; see Figure 2).
{kind=link}
{kind=link}
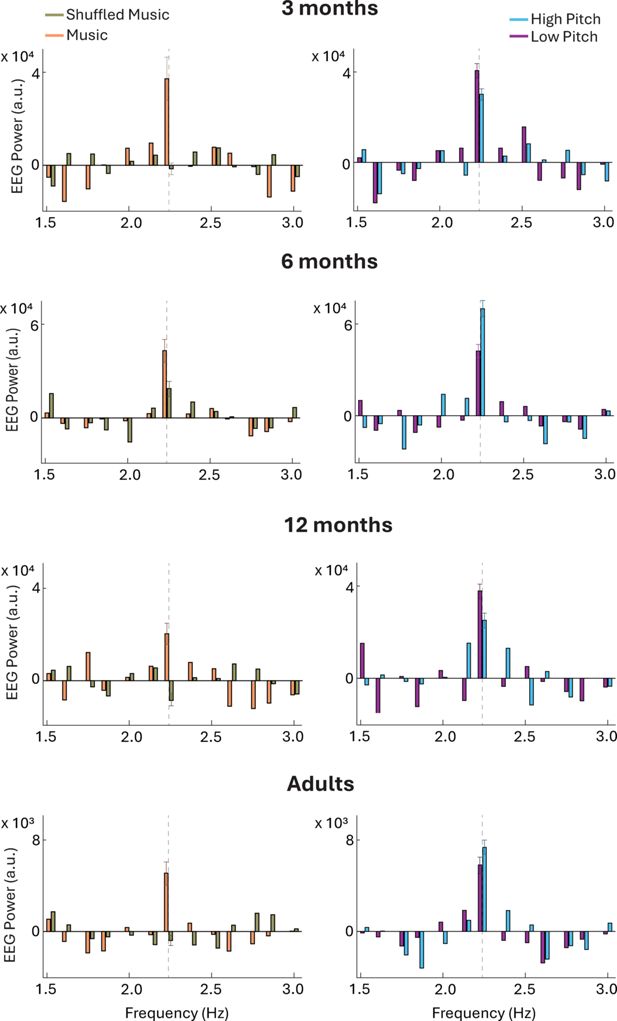
Relative EEG Power (arbitrary units [a.u.], y-axis) of the auditory steady-state responses (ASSR) elicited by music versus shuffled music (orange and khaki, left), and high-pitch versus low-pitch musical stimuli (blue and purple, right), across four participant groups: 3-month-olds (first row), 6-month-olds (second row), 12-month-olds (third row), and adults (fourth row). ASSR power estimates at the frequency (x-axis) matching the musical beat (2.25 Hz, highlighted by vertical dashed lines and including the standard error of the mean) were statistically higher when elicited by music compared to shuffled music across nearly all participant groups (i.e. all but 6-month-olds). High- and low-pitch stimuli evoked similar ASSR (at 2.25 Hz). These results broadly align with the event-related potential (ERP) results (Figure 2) across most infant groups and adults, except for 6-month-old infants for whom differences across conditions were either trending (music vs shuffled) or not significant (high vs low pitch).
Our frequency of interest was 2.25 Hz, matching the musical beat as well as the presentation rate of the majority of the notes (15 out of 16 notes) in the auditory sequences (see methods, section ‘Stimuli’). We used linear mixed models, including power estimates as the dependent variable to contrast the different conditions and age groups (fixed and interaction effects). Participants were modelled as random intercepts. Power estimates were generally higher in response to music as opposed to shuffled music. Model outputs indicated that the power of the ASSR elicited by the music condition was significantly higher than that elicited by the shuffled music condition in 3-month-old infants (F(1,50)=7.82, p=0.007), 12-month-old infants (F(1,52)=12.03, p=0.001) and adults (F(1,50)=13.49, p<0.001); while it only reached a trend for significance in 6-month-old infants (F(1,50)=2.95, p=0.092). ASSR power estimates were not different across high-pitch and low-pitch conditions across all age groups (p>0.240). These results indicated that most groups tracked the periodicity of the musical beat, leading to an enhancement of power in ASSR at a frequency matching the musical beat. This is generally in line with the ERP results showing stronger neural responses to music than shuffled music (even though 6-month-old infants only showed a marginal difference).
Extraction of principal movements and estimation of quantity of movement
Using principal component analysis, we decomposed full-body kinematics into 10 Principal Movements (PMs), explaining 79.7% of the whole kinematic variance. The PMs (depicted in Figure 4) were reminiscent of common infant movements, such as front-back rocking (PM1), side sway (PM2), proto-clapping (PM3), leg kicking (PM4), up-down rocking (PM5), arm pedalling (PM6), feet kicking (PM7), whole body wiggling (PM8), feet shuffling (PM9), and feet pedalling (PM10). Labels were assigned qualitatively, following visual inspection of the PMs.
{kind=link}
{kind=link}
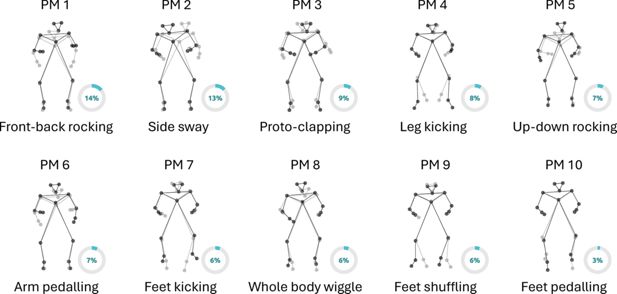
Infants’ principal movements (PMs) are illustrated by showing the two most different body postures (min and max of the PM score, in gray and black, respectively) from the frontal perspective. The reader should interpret the PM as the kinematic displacement necessary to shift from one body posture (gray) to the other (black). Circle diagrams denote the proportion (%) of kinematic variance explained by each PM. Together, the ten PMs account for 79.7% of the total kinematic variance.
Quantity of Movement (QoM) was estimated and compared across PMs, conditions, and groups (Figure 5). QoM estimates were based on first-degree differentiation of the PM time series (see methods). A random intercept was included for each infant. Linear mixed effect modelling yielded a significant interaction between Condition and Age (χ²(2)=16.76, p<0.001) indicating that only 12-month-old infants exhibited higher QoM in response to music compared to shuffled music across all PMs (i.e. when QoM was computed as an average across all PMs). Post-hoc contrasts comparing QoM averages between conditions revealed a highly significant effect (t(69.8)=4.86, p<0.001; see top left panel of Figure 5 for QoM averaged across PMs).
{kind=link}
{kind=link}
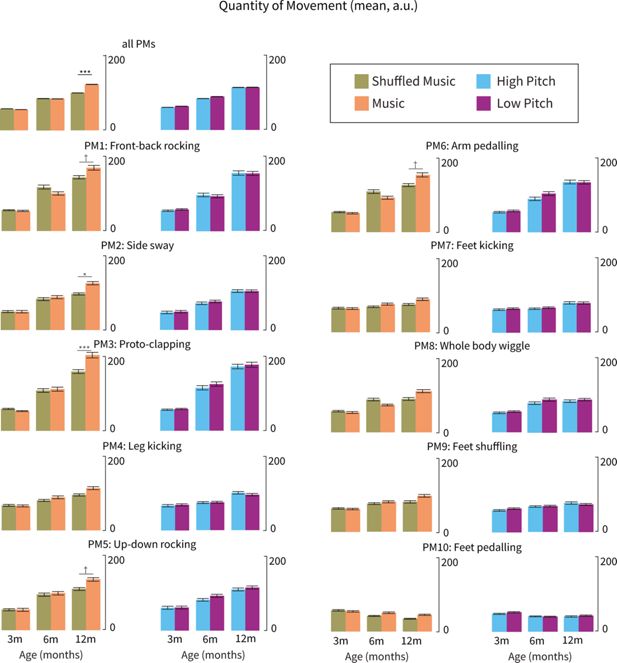
Quantity of movement (QoM; mean, a.u.) elicited by music (orange) versus shuffled music (khaki) and high-pitch (blue) versus low-pitch music (purple) across different age groups (3-month-olds, 6-month-olds, 12-month-olds) and principal movements (PMs). Bar plots indicate the mean and standard error of QoM across different age groups, conditions, and PMs. Only 12-month-old infants showed significantly increased QoM in response to music compared to shuffled music, specifically in PMs involving upper body movements (front-back rocking, side sway, proto-clapping, up-down rocking, and arm pedalling). No significant differences were observed between high- and low-pitch conditions. These results were also replicated in a supplementary analysis assessing differences in variance of (as opposed to mean) QoM (see Appendix 2 for more details). †=p<0.100, =p<0.050, =p<0.010, =p<0.001.
Even though there was no interaction effect between Condition, Age, and PMs, we still ran post-hoc comparisons to gain preliminary evidence about specific PMs driving the above-described effects. Results indicated that differences in 12-month-olds’ QoM in response to music vs shuffled music were mostly driven by movements of the upper body and/or upper limbs. Specifically, front-back rocking (PM1), side sway (PM2), proto-clapping (PM3), up-down rocking (PM5), and arm pedalling (PM6) were linked with significantly higher QoM in response to music as opposed to shuffled music (PMs 1, 2, 3, 5, 6; ps <0.050, corrected using the false-discovery rate). Contrarily, younger infants (3- and 6-month-olds) did not exhibit significantly different QoM in response to music vs shuffled music in any of the PMs (ps >0.123). Furthermore, when comparing infants’ QoM to music at different pitches, we found no significant differences between conditions across PMs and age groups (ps >0.295).
The model also yielded a significant interaction between PMs and Age (χ²(18)=181.575, p<0.001), indicating that QoM generally increased with age but differently across PMs. Specifically, from 3 to 6 months, the PMs associated with a faster increase in QoM were front-back rocking (PM1), proto-clapping (PM3) and arm pedalling (PM6) (t(95.4)=2.34-2.60, p=0.016-0.032). From 3-12 months, front-back rocking (PM1), side sway (PM2), proto-clapping (PM3), up-down rocking (PM5), and arm pedalling (PM6) became more prevalent (t(95.4)=2.69-5.06, p=0.001-0.025). From 6-12 months, proto-clapping (PM3) became even more prevalent (t(95.4)=2.71, p=0.012). Across the first year of life, infants seemed to consistently move their lower body while slowly increasing their capacity for upper-body and whole-body movements while seated.
Movement: Granger causality analysis
Beyond looking at how much infants moved, we further investigated whether the spontaneous occurrence of infant movements could be explained by preceding changes in the intensity of the auditory stimuli. To do so, we used the sound envelope of the auditory stimuli (indexing changes in intensity over time) to predict infant movement velocity (time series representing changes in movement velocity, averaged across all PMs) and vice versa, using Granger causality analysis. Prediction estimates (Granger F-values) were yielded across different time lags, indexing the elapsed time necessary for a change in stimulus intensity to predict a change in movement velocity (i.e. highest F-values represent optimal prediction).
A preliminary analysis (sanity check) showed that musical stimuli predicted subsequent movement velocity better than vice versa across all age groups (Figure 6, top-right corner; F(1)=94.62, p<0.001). To identify the optimal temporal lags predicting sound-driven changes in movement, we conducted bootstrapped t-tests corrected for multiple comparisons across time points. Results (Figure 6, left) showed that musical stimuli were better predictors of movement than shuffled stimuli at 3 months (lags of 160-200 ms, t(40)=3.55-3.98, p<0.001), 6 months (lags of 120-240 ms, t(37)=2.99-4.49, p≤0.001) and 12 months (lags of 160-240 ms, t(31.6)=4.95-5.03, p<0.001). Conversely, there was a drop in prediction at later time lags in all groups (~320-360 ms; t(43.6)=3.88-8.25, p<0.001). Together, these results suggest that infants’ movement was related to intensity changes in the music, but not in the shuffled music.
{kind=link}
{kind=link}
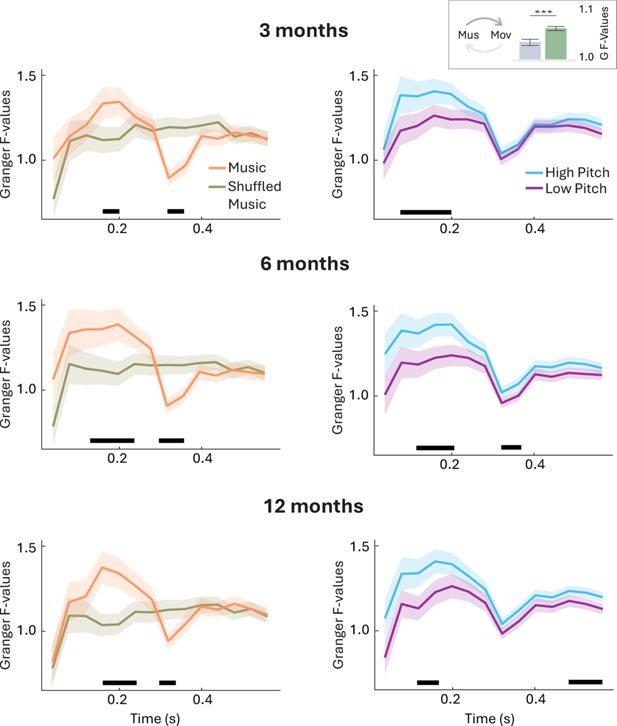
Top-right: A sanity check analysis showed that musical stimuli (i.e. sound envelope) predicted subsequent movement velocity (green) better than vice versa (gray; p<0.001). Left: Movement velocity was better predicted by music (orange) than by shuffled music (khaki), particularly with time lags of 160–200 ms (shaded areas indicate the standard error of the mean ; horizontal black lines underline time ranges associated with a significant difference between conditions). Right: Movement was better predicted by high-pitch music (blue) compared to low-pitch music (purple).
Results associated with the high- and low-pitch conditions yielded similar Granger F-values to the music condition (Figure 6, right). Notably, prediction estimates were generally higher for the high-pitch condition as compared to the low-pitch condition, indicating that high-pitch music was a better predictor of movement than low-pitch music. The difference between these two conditions was significant in one time-window in the 3-month-old infants (80-200 ms, t(39.26)=2.66-3.81, p<0.002), while it encompassed two time windows in the 6-month-olds (120-200 ms, t(45.64)=3.14-4.21, p≤0.004, and 320–360 ms, t(45.7)=3.32-3.82, p≤0.004) and 12-month-olds (120-160 ms, t(40.4)=3.20-3.73, p≤0.002 and 520-600 ms, t(39.9)=3.13-3.41, p<0.004) – perhaps suggesting that this effect grows stronger with age. Finally, Granger Causality statistics stratified by each PM and age group are detailed in the Appendix 2 and Figure 6—figure supplement 1.
Movement: Phase-locked changes and periodicity
The Granger causality analysis indicated that changes in music intensity drove movement in time, especially at a time lag comprised within a 200 ms delay. This result indirectly suggests that changes in music might evoke a phase-locked movement response. If so, we should be able to observe such phase-locked response when epoching movement data to peaks in the amplitude envelope of the auditory stimuli. To test this prediction, we ran supplementary event-related analyses on the movement velocity time series used for the previous analysis. Cluster-based permutation analyses revealed no significant clusters across age groups and conditions (ps >0.050; see Appendix 2, Figure 6—figure supplement 2), even though it should be noted that 12-month-old infants exhibited a movement peak at ~200 ms, which was slightly but not significantly higher in response to music vs shuffled conditions. These results indicated that infants did not consistently exhibit phase-locked movement responses to musical events, such as peaks in the amplitude envelope of the auditory stimuli.
Next, we also examined to what extent spontaneous movements were periodic and how so across conditions. This analysis builds upon the distribution of the highest coefficients yielded by auto-correlation analyses (similar to Zentner and Eerola, 2010). The results indicated that overall, spontaneous movements tended to be periodic, but such periodicity did not match the musical beat and was not different across conditions (see Appendix 2). In other words, movement periodicity did not result in coordination with music, and it was not modulated by whether infants were listening to music vs shuffled music or high- vs low-pitch music (Figure 6—figure supplement 3). This result was further supported by an exploratory analysis in which we identified movement bursts by isolating epochs with high amplitude variation in the time series and excluding segments with low or no variation (c.f. Fujii et al., 2014). Together, these results suggest that while infants might generally exhibit rhythmic movements in response to sounds, the specificity of this behavioural response to music – and its coordination with musical structure – continues to develop beyond 12 months of age.
Discussion
This study examined the development of infants’ neural and movement responses to music during the first year of life using a combination of neural measures, such as ERP and ASSR, alongside quantitative analyses of infants’ movement. These approaches allowed us to explore both sensory and motor components of musicality, shedding light on how infants process and respond to musical stimuli. Below, we discuss (1) the development of neural auditory responses to music, (2) the emergence of music-driven movement patterns, and (3) the sensitivity of both components to pitch across early development.
Neural responses to music: Sensitivity to structured music across all ages
Much research suggests that the infants’ auditory system is sensitive to structural features of music, such as timing and pitch regularities, from early on (Saffran et al., 1999; Thiessen and Saffran, 2009; see Nguyen et al., 2023a for a review). However, the developmental trajectory of such musical sensitivity across the first year of life is still underexplored. Here, we provide a detailed characterization of the progressive maturation of auditory evoked potentials, neurophysiological measures that generally capture rapid changes in the sensory environment, such as the onset of musical notes (Kushnerenko et al., 2002; Somervail et al., 2021). Specifically, we observed ERPs with progressively shorter latency and larger amplitude throughout the first year of life. While these findings likely reflect a general maturation of sensory systems, they also highlight how the evoked potentials elicited by musical stimuli were enhanced in amplitude compared to those elicited by shuffled (i.e. structure-free) stimuli, an effect consistently observed across all age groups. This is in line with the idea that sensitivity to musical regularities emerges early and persists throughout infancy (Trevarthen, 1999). Indeed, sensitivity to simple rhythmic or pitch regularities has been observed in infants at various ages (Cirelli et al., 2016; Edalati et al., 2023; Flaten et al., 2022; Háden et al., 2009; Háden et al., 2024; Stefanics et al., 2009; Winkler et al., 2009), and, in the case of rhythm, even in premature neonates (Saadatmehr et al., 2025). Such sensitivity may be rooted in general auditory predictive processing, whereby the brain extracts regularities from past observations to generate and update predictions about incoming sensory information (Friston, 2010; Köster et al., 2020; Vuust and Witek, 2014). Ecologically valid music, such as the refrains used here, naturally incorporates rhythmic and pitch regularities that could trigger the generation of predictions. This process was markedly dampened or interrupted by shuffled music (Bianco et al., 2024; Bianco et al., 2025; Lense et al., 2022), a finding that could be interpreted as evidence of disengagement from such highly unpredictable sequences (Dayan et al., 2000; Esber and Haselgrove, 2011; Kidd et al., 2014).
[Kidd e